Candidate Genes for Freezing and Drought Tolerance Selected on the Basis of Proteome Analysis in Doubled Haploid Lines of Barley

 ,
,  , ,
, ,  ,
,  ,
,
Abstract
1. Introduction
2. Results
2.1. Candidate Genes Selection
- 1.
- Elongation factor 1 alpha (EF1A) coding gene (primers designed for the consensus mRNA sequence of: KP293845.1 and KP293846.1);
- 2.
- Ferredoxin-NADP reductase coding gene (mRNA sequence ID: AK368450.1);
- 3.
- Gene encoding 14-3-3a protein (mRNA sequence ID: X62388.1);
- 4.
- Gene encoding β-fructofuranosidase (primers designed on the basis of a consensus mRNA sequence for six splicing variants of MF443751);
- 5.
- CBF 4B (primers designed for the consensus mRNA sequence of: DQ480160.1:7462-8139, DQ445234.1:4551-5228, AY785853.1:86-763 and AY785848.1:84-761);
- 6.
- CBF2A (primers designed for the consensus mRNA sequence of: DQ480160.1:14050-14715, GU461589.1:13-678, GU461588.1:15-680, GU461587.1:21-686 and AY785840.1:13-678).
- 1.
- Transketolase gene (mRNA sequence ID: AK362454.1:1-2041);
- 2.
- Gene encoding periplasmic serine protease (mRNA sequence ID: AK355966.1:49-1332);
- 3.
- Triosephosphate isomerase gene (mRNA sequence ID: AK375585.1:86-847);
- 4.
- A protein-coding gene with a co-chaperonin region (GroEs) (primers designed for the consensus mRNA sequence of: AK369605.1:156-911 and AK362060.1:215-970);
- 5.
- Gene encoding pfam14200—ricin-type beta-trefoil lectin domain-like protein (mRNA sequence ID: AK372454.1);
- 6.
- Actin gene (mRNA sequence ID: AY145451).
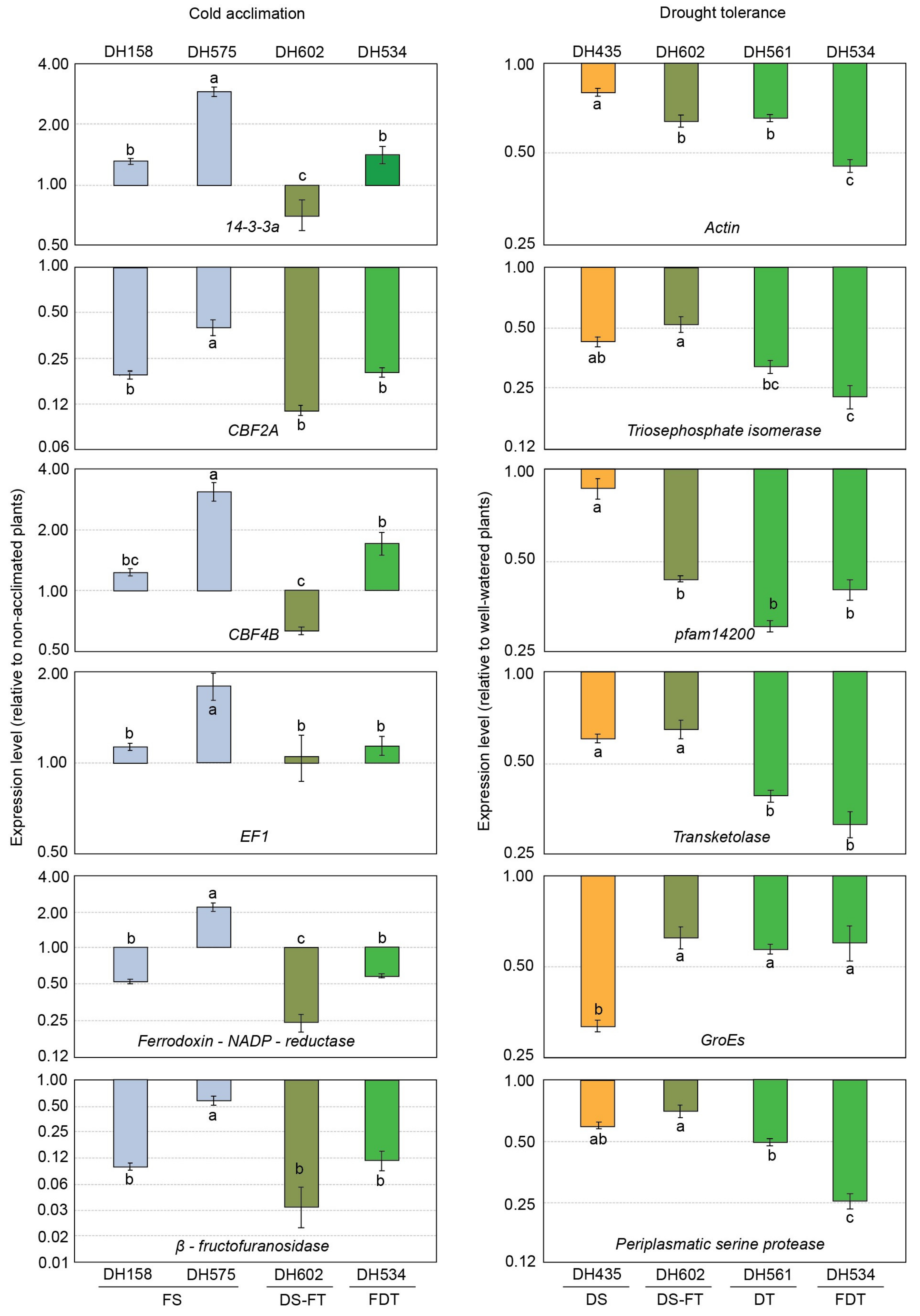
2.2. Changes in Expression of Selected Genes under Abiotic Stress Conditions
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Plant Growth and Stress Treatments
4.3. RNA Isolation and Reverse Transcription
4.4. Candidate Genes Selection
4.5. RT-qPCR
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MTL | Mendelian Trait Loci |
| QTL | Quantitative Trait Loci |
| DH | double haploid |
| EF1a | elongation factor 1 alpha |
| RT qPCR | quantitative (real-time) PCR performed on cDNA template |
| NA | not acclimated to cold |
| CA | cold-acclimated |
| C | control (well-watered plants) |
| D | drought-treated plants |
| ABA | abscisic acid |
| CRT | C-repeat |
| Hsp | heat shock protein |
| SWC | soil water content |
| CDS | coding DNA sequence |
| ADP | ADP-ribosylation factor 1-like protein |
| sAMD | S-adenosylmethionine decarboxylase |
| CT | threshold cycle |
References
- Tricker, P.J.; ElHabti, A.; Schmidt, J.; Fleury, D. The physiological and genetic basis of combined drought and heat tolerance in wheat. J. Exp. Bot. 2018, 69, 3195–3210. [Google Scholar] [CrossRef]
- Sheoran, S.; Malik, R.; Narwal, S.; Tyagi, B.S.; Mittal, V.; Kharub, A.S.; Tiwari, V.; Sharma, I. Genetic and molecular dissection of drought tolerance in wheat and barley. J. Wheat Res. 2016, 7, 1–13. [Google Scholar]
- Gusta, L.V.; Wisniewski, M. Understanding plant cold hardiness: An opinion. Physiol. Plant. 2013, 147, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Byrne, P.F.; McMullen, M.D. Defining genes for agricultural traits: QTL analysis and the candidate gene approach. Probe 1996, 7, 24–27. [Google Scholar]
- Pflieger, S.; Lefebvre, V.; Causse, M. The candidate gene approach in plant genetics: A review. Mol. Breed. 2001, 7, 275–291. [Google Scholar] [CrossRef]
- Zhu, M.; Zhao, S. Candidate gene identification approach: Progress and challenges. Int. J. Biol. Sci. 2007, 3, 420. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, C.; Kia, D.A.; Vandrovcova, J.; Hardy, J.; Wood, N.W.; Lewis, P.A.; Ferrari, R. Genome, transcriptome and proteome: The rise of omics data and their integration in biomedical sciences. Brief. Bioinform. 2018, 19, 286–302. [Google Scholar] [CrossRef] [PubMed]
- Gołębiowska-Pikania, G.; Kopeć, P.; Surówka, E.; Krzewska, M.; Dubas, E.; Nowicka, A.; Rapacz, M.; Wójcik-Jagła, M.; Malaga, S.; Żur, I. Changes in protein abundance and activity involved in freezing tolerance acquisition in winter barley (Hordeum vulgare L.). J. Proteom. 2017, 169, 58–72. [Google Scholar] [CrossRef]
- Gołębiowska-Pikania, G.; Kopeć, P.; Surówka, E.; Janowiak, F.; Krzewska, M.; Dubas, E.; Nowicka, A.; Kasprzyk, J.; Ostrowska, A.; Malaga, S.; et al. Changes in protein abundance and activity induced by drought during generative development of winter barley (Hordeum vulgare L.). J. Proteom. 2017, 169, 73–86. [Google Scholar] [CrossRef]
- Malaga, S.; Janeczko, A.; Janowiak, F.; Waligórski, P.; Oklestkova, J.; Dubas, E.; Krzewska, M.; Nowicka, A.; Surówka, E.; Rapacz, M.; et al. Involvement of homocastasterone, salicylic and abscisic acids in the regulation of drought and freezing tolerance in doubled haploid lines of winter barley. Plant Growth Regul. 2019, 90, 173–188. [Google Scholar] [CrossRef]
- Busconi, M.; Dal Bosco, C.; Crosatti, C.; Baldi, P.; Marè, C.; Grossi, M.; Mastrangelo, A.M.; Rizza, F.; Cattivelli, L.; Stanca, A.M. The cold-regulated genes are involved in the physiological response of barley to cold environment. Icel. Agric. Sci. 2001, 14, 17–27. [Google Scholar]
- Ceccarelli, S. Drought and Drought Resistance. In Encyclopedia of Biotechnology in Agriculture and Food, 1st ed.; Heldman, D.R., Wheeler, M.B., Hoover, D.G., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 205–207. [Google Scholar]
- Yan, G.; Liu, H.; Wang, H.; Lu, Z.; Wang, Y.; Mullan, D.; Hamblin, J.; Liu, C. Accelerated Generation of Selfed Pure Line Plants for Gene Identification and Crop Breeding. Front. Plant Sci. 2017, 8, 1786. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, M.W.; Gąsior, D.; Leśniewska-Bocianowska, A.; Zwierzykowski, Z.; Rapacz, M. Androgenesis as a means of dissecting complex genetic and physiological controls: Selecting useful gene combinations for breeding freezing tolerant grasses. Euphytica 2007, 158, 337–345. [Google Scholar] [CrossRef]
- Yonezawa, K.; Ishii, T. Marker-based selection as a tool for enhancing the efficiency of two conventional breeding methods of self-fertilizing crop plants, the generation-accelerated bulk breeding and doubled haploid breeding. Breed. Sci. 2005, 55, 397–407. [Google Scholar] [CrossRef]
- Malaga, S.; Krzewska, M.; Nowicka, A.; Dubas, E.; Ostrowska, A.; Wójcik-Jagła, M.; Hura, T.; Rapacz, M.; Żur, I. Increasing tolerance to abiotic stresses by haplodiploidization in Hordeum vulgare. In Plant Functioning under Environmental Stress, 1st ed.; Grzesiak, M., Rzepka, A., Hura, T., Grzesiak, S., Eds.; The F. Górski Institute of Plant Physiology, Polish Academy of Sciences: Kraków, Poland, 2016; pp. 37–46. [Google Scholar]
- Fiust, A.; Rapacz, M. Downregulation of three novel candidate genes is important for freezing tolerance of field and laboratory cold acclimated barley. J. Plant Physiol. 2020, 244, 153049. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhou, G.; Shimizu, H. Plant responses to drought and rewatering. Plant Signal. Behav. 2010, 5, 649–654. [Google Scholar] [CrossRef]
- Francia, E.; Barabaschi, D.; Tondelli, A.; Laidò, G.; Rizza, F.; Stanca, A.M.; Busconi, M.; Fogher, C.; Stockinger, E.J.; Pecchioni, N. Fine mapping of a HvCBF gene cluster at the frost resistance locus Fr-H2 in barley. Theor. Appl. Genet. 2007, 115, 1083–1091. [Google Scholar] [CrossRef]
- Skinner, J.S.; von Zitzewitz, J.; Szűcs, P.; Marquez-Cedillo, L.; Filichkin, T.; Amundsen, K.; Stockinger, E.J.; Thomashow, M.F.; Chen, T.H.H.; Hayes, P.M. Structural, functional, and phylogenetic characterization of a large CBF gene family in barley. Plant Mol. Biol. 2005, 59, 533–551. [Google Scholar] [CrossRef]
- Faccioli, P.; Ciceri, G.P.; Provero, P.; Stanca, A.M.; Morcia, C.; Terzi, V. A combined strategy of ‘‘in silico’’ transcriptome analysis and web search engine optimization allows an agile identification of reference genes suitable for normalization in gene expression studies. Plant Mol. Biol. 2007, 63, 679–688. [Google Scholar] [CrossRef]
- Rapacz, M.; Stępień, A.; Skorupa, K. Internal standards for quantitative RT-PCR studies of gene expression under drought treatment in barley (Hordeum vulgare L.): The effects of developmental stage and leaf age. Acta Physiol. Plant. 2012, 34, 1723–1733. [Google Scholar] [CrossRef]
- Śniegowska-Świerk, K.; Dubas, E.; Rapacz, M. Actin microfilaments are involved in the regulation of HVA1 transcript accumulation in drought-treated barley leaves. J. Plant Physiol. 2016, 193, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Esch, L.; Schaffrath, U. An Update on Jacalin-Like Lectins and Their Role in Plant Defense. Int. J. Mol. Sci. 2017, 18, 1592. [Google Scholar] [CrossRef] [PubMed]
- Lanet, E.; Delannoy, E.; Sormani, R.; Floris, M.; Brodersen, P.; Crété, P.; Voinnet, O.; Robaglia, C. Biochemical evidence for translational repression by Arabidopsis microRNAs. Plant Cell 2009, 21, 1762–1768. [Google Scholar] [CrossRef] [PubMed]
- Iwakawa, H.; Tomari, Y. Molecular insights into microRNA-mediated translational repression in plants. Mol. Cell 2013, 52, 591–601. [Google Scholar] [CrossRef]
- Hochstrasser, M. Ubiquitin-dependent protein degradation. Annu. Rev. Genet. 1996, 30, 405–439. [Google Scholar] [CrossRef]
- Callis, J.; Vierstra, R.D. Protein degradation in signaling. Curr. Opin. Plant Biol. 2000, 3, 381–386. [Google Scholar] [CrossRef]
- Gilmour, S.J.; Zarka, D.G.; Stockinger, E.J.; Salazar, M.P.; Houghton, J.M.; Thomashow, M.F. Low temperature regulation of the Arabidopsis CBF family of AP2 transcriptional activators as an early step in cold-induced COR gene expression. Plant J. 1998, 16, 433–442. [Google Scholar] [CrossRef]
- Salvador, M.L.; Klein, U.; Bogorad, L. Light-regulated and endogenous fluctuations of chloroplast transcript levels in Chlamydomonas. Regulation by transcription and RNA degradation. Plant J. 1993, 3, 213–219. [Google Scholar] [CrossRef]
- Shu, Y.; Hong-Hui, L. Transcription, translation, degradation, and circadian clock. Biochem. Biophys. Res. Commun. 2004, 321, 1–6. [Google Scholar] [CrossRef]
- Jacquard, C.; Wojnarowiez, G.; Clément, C. Anther culture in barley. In Doubled Haploid Production in Crop Plants, 1st ed.; Małuszynski, M., Kasha, K.J., Forster, B.P., Szarejko, I., Eds.; Kluwer Academic Publisher: Dordrecht, The Netherlands, 2003; pp. 21–28. [Google Scholar]
- Cistué, I.; Vallés, M.P.; Echávarri, B.; Sanz, J.M.; Castillo, A. Barley anther culture. In Doubled Haploid Production in Crop Plants, 1st ed.; Małuszynski, M., Kasha, K.J., Forster, B.P., Szarejko, I., Eds.; Kluwer Academic Publisher: Dordrecht, The Netherlands, 2003; pp. 29–34. [Google Scholar]
- Rapacz, M.; Sasal, M.; Gut, M. Chlorophyll fluorescence-based studies of frost damage and the tolerance for cold-induced photoinhibition in freezing tolerance analysis of triticale (xTriticosecale Wittmack). J. Agron. Crop. Sci. 2011, 197, 378–389. [Google Scholar] [CrossRef]
- Hura, T.; Tyrka, M.; Hura, K.; Ostrowska, A.; Dziurka, K. QTLs for cell wall-bound phenolics in relation to the photosynthetic apparatus activity and leaf water status under drought stress at different growth stages of triticale. Mol. Genet. Genom. 2017, 292, 415–433. [Google Scholar] [CrossRef] [PubMed]
- Lancashire, P.D.; Bleiholder, H.; Boom, T.V.D.; Langelüddeke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A uniform decimal code for growth stages of crops and weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- NCBI. Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=tblastn&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome (accessed on 20 July 2018).
- EMBL-EBI. Lalign. Available online: https://www.ebi.ac.uk/Tools/psa/lalign/ (accessed on 21 July 2018).
- EMBL-EBI. Clustal-omega. Available online: https://www.ebi.ac.uk/Tools/msa/clustalo/ (accessed on 21 July 2018).
- Primer3Plus. Available online: https://primer3plus.com/cgi-bin/dev/primer3plus.cgi (accessed on 22 July 2018).
- Bustin, S.A. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J. Mol. Endocrinol. 2000, 25, 169–193. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Protein ID | Query Cover | E Value | Percent Identity | Coding or Genomic Sequence ID | CDS Annotation |
|---|---|---|---|---|---|
| KQJ82088.1 | 100% | 0.0 | 95.98% | KY636108.1 | - |
| EPS62279.1 | 54% | 1 × 10−35 | 30.96% | JN818424.1 | cytochrome P450 |
| EMT12632.1 | 100% | 4 × 10−180 | 94.51% | AK253057.1 | - |
| 100% | 4 × 10−162 | 84.71% | AK368450.1 | CDS for predicted protein (ferredoxin--NADP(+) reductase) | |
| EMT33607.1 | 100% | 0.0 | 94.68% | AK250604.1 | - |
| 95% | 1 × 10−33 | 29.12% | JN107538.1 | H. brevisubulatum elongation factor 1 alpha | |
| 95% | 7 × 10−33 | 27.60% | KP293846.1 | H. vulgare eukaryotic elongation factor 1 alpha | |
| KQK13608.1 | 92% | 1 × 10−173 | 100.00% | X62388.1 | H. vulgare 14-3-3 protein homologue (14-3-3a) |
| 91% | 5 × 10−152 | 86.89% | DQ295786.1 | H. vulgare subsp. vulgare 14-3-3E | |
| 90% | 1 × 10−151 | 88.19% | X93170.1 | H. vulgare Hv14-3-3b | |
| 90% | 4 × 10−151 | 87.45% | Y14200.1 | H. vulgare 14-3-3 protein (Hv1433c) | |
| 91% | 1 × 10−131 | 78.75% | DQ295785.1 | H. vulgare subsp. vulgare 14-3-3D | |
| XP_013654063.1 | 72% | 2 × 10−34 | 53.64% | MF443751.1 | putative beta-fructofuranosidase |
| 73% | 2 × 10−31 | 48.65% | AY266442.1 | Mla6-2 gene, complete cds | |
| 79% | 1 × 10−21 | 41.60% | AF509748.1 | H. vulgare subsp. vulgare Morex barley stem rust resistance protein (Rpg1) | |
| 90% | 1 × 10−37 | 48.55% | DQ480160.1 | putative glutaredoxin protein, CBF4B and CBF2A | |
| 90% | 1 × 10−37 | 48.55% | DQ445234.1 | putative glutaredoxin protein, CBF4B and CBF2A | |
| BAJ98295.1 | 93% | 0.0 | 95.52% | AK368450.1 | CDS for predicted protein (ferredoxin−-NADP(+) reductase) |
| 93% | 0.0 | 95.52% | AK367092.1 | CDS for predicted protein (ferredoxin--NADP(+) reductase) | |
| 91% | 0.0 | 77.81% | AK253057.1 | - | |
| BAJ93658.1 | 97% | 0.0 | 100.00% | AK362454.1 | partial CDS for predicted protein (transketolase) |
| BAK06780.1 | 100% | 7 × 10−159 | 89.33% | AK375585.1 | - |
| 100% | 3 × 10−157 | 88.93% | U83414.1 | H. vulgare cytosolic triosephosphate isomerase | |
| KXG22555.1 | 76% | 0.0 | 93.11% | AK355966.1 | periplasmic serine protease, S1-C subfamily |
| 72% | 9 × 10−85 | 50.00% | AK362697.1 | periplasmic serine protease, S1-C subfamily | |
| EMT10427.1 | 100% | 7 × 10−163 | 81.13% | AK369605.1 | co-chaperonin GroES |
| 100% | 1 × 10−161 | 81.46% | AK362060.1 | co-chaperonin GroES | |
| AAX12161.1 | 100% | 5 × 10−58 | 91.75% | AY145451.1 | Hordeum vulgare actin mRNA, complete cds |
| BAK03652.1 | 47% | 1 × 10−74 | 68.18% | AK372188.1 | region ricin-type beta-trefoil lectin domain-like; pfam14200 |
| 91% | 3 × 10−153 | 76.09% | AK372454.1 | region ricin-type beta-trefoil lectin domain-like; pfam14200 |
| Gene | Forward/Reverse | Primer Sequence (5′-3′) |
|---|---|---|
| Elongation factor 1 alpha | forward | TGCCACTTACCCTCCTCTTG |
| reverse | TTCTTCTCCACGCCCTTGAT | |
| Ferredoxin-NADP reductase | forward | GGCGGGAGAGAAGATGTACA |
| reverse | TCAGCCCACACATGTACACA | |
| 14-3-3a protein gene | forward | TTGGGCTTGCACTCAACTTC |
| reverse | GGGAGTCCAGCTCAGCAATA | |
| β-fructofuranosidase | forward | CCGACCCTTTGCTCATCAAC |
| reverse | GGGTCCCTGAAGTCCTTCTC | |
| CBF4B | forward | TTCTCTGGCCTCGCTCTTTC |
| reverse | CGCCGCTCTGTTTTACATCT | |
| CBF2A | forward | ATGATGCGTGCCTCAACTTC |
| reverse | GACGGCGTCCTTGATCTCTT | |
| Transketolase | forward | TTGACGAAGGAGGGGAAGAC |
| reverse | GGTAGAGCCAGCTTCAATGC | |
| Periplasmic serine protease | forward | AAGCGCAAGTTGTCGGATTT |
| reverse | CCAGTAGGTCTGCTGACACA | |
| Triosephosphate isomerase | forward | AACTCTGAACGCTGGACAGA |
| reverse | GCAGTTCTGAGCAGCAACTT | |
| GroEs | forward | AGAGGAAACTGCTGGTGGTT |
| reverse | CTGCTTCCAGGAGTGATCGA | |
| Pfam14200 | forward | GTCCCACCCTGTTCTTCTGA |
| reverse | CCATGGAGCGCATCAAAGTT | |
| Actin | forward | CGACAATGGAACCGGAATG |
| reverse | CCCTTGGCGCATCATCTC | |
| ADP-ribosylation factor 1-like protein | forward | CGTGACGCTGTGTTGCTTGT |
| reverse | CCGCATTCATCGCATTAGG | |
| S-adenosylmethionine decarboxylase | forward | TCGGCTACAGCATTGAAGACG |
| reverse | CCAAAAACGATATCAGGATGGC | |
| GADPH | forward | TTCGGCGAGAAGCCAGTTA |
| reverse | CCTCACCCCACGGGATCT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wójcik-Jagła, M.; Rapacz, M.; Dubas, E.; Krzewska, M.; Kopeć, P.; Nowicka, A.; Ostrowska, A.; Malaga, S.; Żur, I. Candidate Genes for Freezing and Drought Tolerance Selected on the Basis of Proteome Analysis in Doubled Haploid Lines of Barley. Int. J. Mol. Sci. 2020, 21, 2062. https://doi.org/10.3390/ijms21062062
Wójcik-Jagła M, Rapacz M, Dubas E, Krzewska M, Kopeć P, Nowicka A, Ostrowska A, Malaga S, Żur I. Candidate Genes for Freezing and Drought Tolerance Selected on the Basis of Proteome Analysis in Doubled Haploid Lines of Barley. International Journal of Molecular Sciences. 2020; 21(6):2062. https://doi.org/10.3390/ijms21062062
Chicago/Turabian StyleWójcik-Jagła, Magdalena, Marcin Rapacz, Ewa Dubas, Monika Krzewska, Przemysław Kopeć, Anna Nowicka, Agnieszka Ostrowska, Sabina Malaga, and Iwona Żur. 2020. "Candidate Genes for Freezing and Drought Tolerance Selected on the Basis of Proteome Analysis in Doubled Haploid Lines of Barley" International Journal of Molecular Sciences 21, no. 6: 2062. https://doi.org/10.3390/ijms21062062
APA StyleWójcik-Jagła, M., Rapacz, M., Dubas, E., Krzewska, M., Kopeć, P., Nowicka, A., Ostrowska, A., Malaga, S., & Żur, I. (2020). Candidate Genes for Freezing and Drought Tolerance Selected on the Basis of Proteome Analysis in Doubled Haploid Lines of Barley. International Journal of Molecular Sciences, 21(6), 2062. https://doi.org/10.3390/ijms21062062